Begerlavslekta Cladonia er ei artsrik lavslekt med omtrent 90 arter i Norge. Slekta omfatter to morfologiske grupper som tidligere har vært oppfatta som underslekter, reinlavene Cladina og de 'ekte begerlavene' Cladonia.
Innhold

Bunnsjikt i furuskog dominert av reinlavarter. Lys reinlav Cladonia arbuscula, grå reinlav Cladonia rangiferina, kvitkrull Cladonia stellaris og pigglav Cladonia uncialis.
Bygning
De fleste begerlavene har et todelt tallus som består av en horisontaldel kalt basalskjell og en vertikaldel som kalles et podetium. Basalskjellene er relativt små, noen få mm til noen cm lange, og sitter mer eller mindre tett sammen. Det er vanligvis tydelig forskjell mellom oversiden som er barkkledd og undersiden som er uten bark. Basalskjellene kan være mer eller mindre oppdelt og hos noen arter produseres soredier eller pyknidier på dem. Hos f.eks. polsterlav Cladonia strepsilis og kalkpolster Cladonia symphycarpa er basalskjellene helt dominerende og sitter tett sammen i en puteaktig vekstform. Hos de fleste artene som normalt danner podetier er basalskjellene vedvarende gjennom hele livsløpet, men hos noen arter blir de tilbakedannet. Reinlavene mangler basalskjell.
Podetiene vokser ut fra basaldelen og kan være utforma på mange ulike vis. De kan være begerforma, sylforma, stavforma, enkelt gaffelgreina eller rikt buskforma med opprette spisser, eller ha ensidig til allsidig bøyde greinspisser. Bark på podetiene forekommer hos de fleste artene, men ikke hos reinlavene. Dette gir reinlavene en filtaktig overflatestruktur (må studeres med lupe!) fordi marghyfene peker i alle mulige retninger. Hos barkkledde podetier vil hyfene være organisert i en parallell struktur som gir en glattere overflate. En del sorediøse arter har også lite og flekkvis bark. Podetiene kan hos noen arter være forsynt med utstående skjell eller de kan ha barkkledde gryn eller soredier. Begerforma podetier kan være lukka eller åpne hos ulike arter og de kan danne nye begre fra kanten eller fra sentrum av det gamle begeret, i flere etasjer i noen tilfeller. Hos buskforma arter kan greinvinklene være åpne eller lukka. Hos noen arter kan podetiene være forsynt med lengdefurer eller de kan være mer eller mindre oppsprukket i lengderetningen. Algekomponenten hos begerlavene er hovedsakelig encellete grønnalger i slekta Asterochloris.
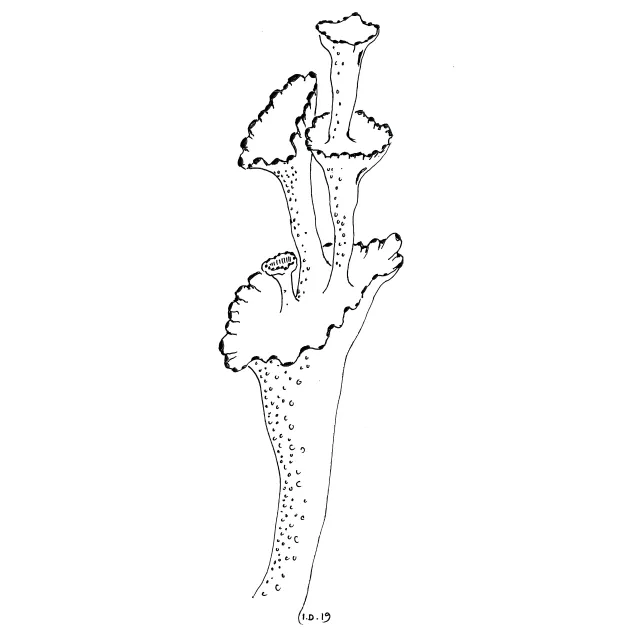
Begerforma podetium som danner nye begre fra midten av det gamle. Mørke punkter langs begerkanten er pyknidier.
Begerforma podetium som danner nye begre fra kanten av det gamle.
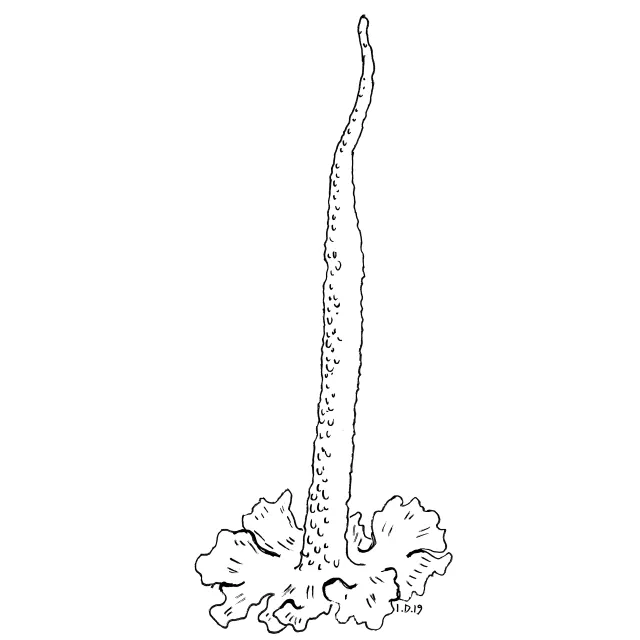
Basalskjell og sylforma podetium med soredier.
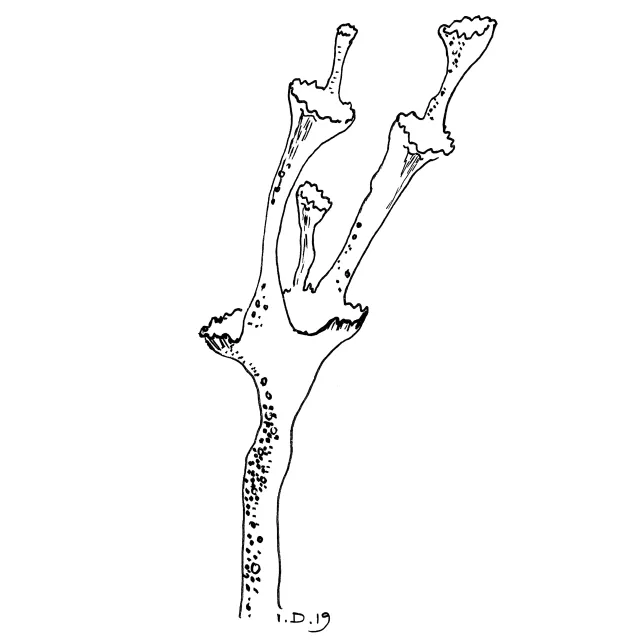
Gaffelgreina podetium med utstående skjell og åpne greinvinkler.
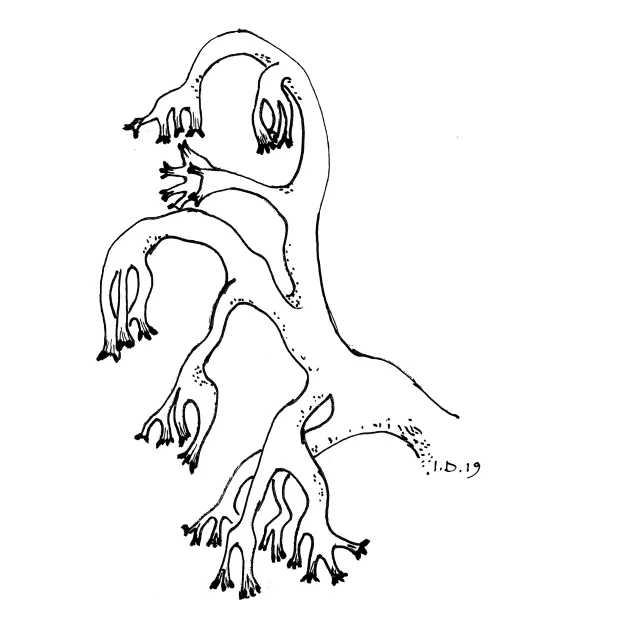
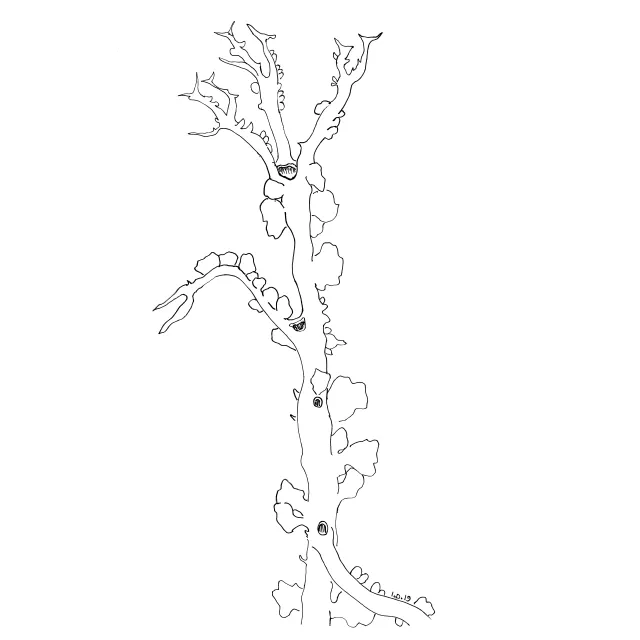
Rikt greina podetium av grå reinlav med ensidig bøyde greinspisser. Mørke greinspisser er pyknidier.
I denne framstillinga er slekta delt i grupper. Disse er basert på morfologiske karakterer og farger på fruktlegemer, og trenger ikke ha noe med slektskap å gjøre. Grupperinga må derfor betraktes som en praktisk måte å dele inn en stor slekt på og det kan argumenters for andre måter å organisere dette på.
Reproduksjon
Reproduksjonen hos begerlavene kan skje på ulike vis. Hos soppkomponenten kan den skje gjennom en kjønna prosess ved dannelse av fruktlegemer (apothecier), eller ukjønna ved produksjon av konidier i små organ kalt pyknidier. Reproduksjonen kan også skje vegetativt gjennom fragmentering av podetier og basalskjell, eller ved dannelse av små barkløse korn, såkalte soredier, eller barkkledde gryn på overflata av podetiene. Reproduksjon med soredier er utbredt hos mange arter, særlig hos arter med beger- eller sylforma podetier.
Fruktlegemene, hvor den kjønna prosessen foregår, er plassert i enden av podetiene og kan være av tre ulike fargetyper, skarlagensrøde, bleikt kjøttfarga (som kyllingkjøtt) eller brune. Sporene er enkle, ellipsoide og fargeløse. Pyknidier er bittesmå flaskeforma beholdere som produserer små ukjønna sporer som kalles konidier. Pyknidiene kan observeres som små stifter med en pore i enden og er plassert på begerkanten eller innsenka i spissen av greiner, sjelden også på oversiden eller i kanten av basalskjellene. De har gjerne samme farge som fruktlegemene for den aktuelle arten dvs. brune eller røde. Konidiene er sigdforma og gjerne mindre enn sporene.

Reinlavsamfunn i kysthei, Flatanger. Lys reinlav Cladonia arbuscula, gaffelreinlav Cladonia ciliata, grå reinlav Cladonia rangiferina og svartfotreinlav Cladonia stygia.
Økologi
Generelt er de fleste artene i begerlavslekta pionerarter på jord, sand og grus, torv, råtnende ved og på basis av trær. De opptrer gjerne tidlig i suksesjoner etter hogst, brann og andre forstyrrelser. Reinlavene kan imidlertid danne relativt stabile samfunn på næringsfattig mark i kontinentale strøk som f.eks. i lavfuruskog, lavbjørkeskog og i taiga- og tundraområder. Ellers opptrer begerlaver og reinlav vanlig i blokkmarker, på berg med tynt jorddekke, i kystheier, alpine områder, i relativt lysåpne skoger, på tuer i ombrotrofe myrer samt på strandberg og i sanddyner. Gode lokaliteter er også hogstflater og yngre plantefelt hvor stubbene kan være helt dominert av begerlaver noen år før skogbestandet fortettes og lysforholdene blir så redusert at mosene overtar. Vegskjæringer og andre steder med forstyrrelser i bunnsjiktet kan også være gode habitat en kortere periode.

Blokkmark i kontinental furuskog med kompakte matter av grå reinlav.
Begerlavene kan ha mange ulike lavsyrer. Dette er organiske forbindelser som produseres av soppkomponenten. Noen lavsyrer fungerer som antibeitestoffer, noen kan beskytte mot UV-lys og noen er pigmenter uten kjent funksjon. Et vanlig pigment er usninsyre som gir en bleikgul farge på tallus. Usninsyre har vist seg å være bakteriedrepende og har vært prøvd ut i medisinen som antibiotikum. I mange tilfeller er det nødvendig å finne ut hvilke stoffer et eksemplar inneholder for å kunne bestemme arten sikkert. Noen stoffer fluorescerer sterkt under UV-lys og kan påvises ved hjelp av en UV-lampe. For eksempel squamatsyre og perlatolsyre gir en kraftig blåhvit fluorescens under UV-lys. Til bestemmelse av en del andre lavsyrer brukes gjerne noen enkle fargereagenser. De vanligste er vanlig klorin (C), 10 % løsning av kalilut (K) og stoffet parafenylendiamin (PD) løst i etanol. Et vanlig stoff hos mange begerlaver er fumarprotocetrarsyre. Når en liten dråpe PD avsettes på en lav som inneholder denne syren oppstår en rød flekk. Vi sier da at laven reagerer PD+ rød. Andre eksempler er thamnolsyre som blir K+ kraftig gul, psoromsyre som blir PD+ svovelgul og strepsilin som blir C+ irrgrønn. Noen lavsyrer kan bare påvises ved hjelp av mer kompliserte metoder som tynnsjiktkromatografi (TLC). Eksempler på slike lavsyrer som forekommer hos flere begerlaver er barbatinsyre, terpenet zeorin og fettsyren rangiformsyre.

Reinlavsamfunn i mosaikk med heigråmose Racomitrium lanuginosum i kysthei, Flatanger.
Mange begerlaver har stor intern variasjon noe som gjør at nøkling av arter kan være svært vanskelig. Nøklene her er basert på typisk og godt utvikla materiale. En vil oppleve at eksemplarer fra uvanlige habitat eller dårlig utvikla materiale vil være vanskelig, eller umulig å bestemme sikkert. Særlig gjelder dette innenfor enkelte artskomplekser. Til sikker bestemmelse vil det i mange tilfeller være nødvendig med en UV-lampe og de tre fargereagensene C, K og PD nevnt ovenfor. UV-lamper brukt av frimerkesamlere kan benyttes, men det beste vil være lamper som har både kortbølget og langbølget UV-lys.

Begerlavsamfunn på svært nedbrutt furustubbe i Varnvassdalen, Hattfjelldal. Dominerende arter her er blomsterlav Cladonia bellidiflora og fingerbeger Cladonia digitata. I øvre del vises også litt meltraktlav Cladonia cenotea og fausklav Cladonia sulphurina.

Fattig furuskog på grunnlendt mark. I forgrunnen er bunnsjiktet dominert av reinlavsamfunn. I bakkant har bunnsjiktet tykkere humusdekke og er dominert av lyngarter med bare flekkvis forekomst av reinlav.
Ahti T, Stenroos S og Moberg R (eds.) (2013). Cladoniaceae. Nordic Lichen Flora, Vol. 5. Nordic Lichen Society, Uppsala.
Holien H og Tønsberg T (2008). Norsk lavflora. 2. utgave. Tapir akademisk forlag, Trondheim.
Krog H, Østhagen H og Tønsberg T (1994). Lavflora. Norske busk- og bladlav. 2. utgave. Universitetsforlaget, Oslo.
Moberg R og Hultengren S (2016). Lavar. En fältguide. Naturcentrum AB, Stenungsund.
Stenroos S, Velmala S, Pykälä J og Ahti T (2016). Lichens of Finland. Norrlinia 30: 1-896.
Søchting U (2017). Lav i klit og hede. De danske rensdyr- og bægerlaver og deres følgearter. Biologisk forening for Nordvestjylland, København.
Om denne siden
Publisert: 23. januar 2017Artshåndbok
Beskrivelsen er en del av en artshåndbok.

Fra artshåndboken
Siden siteres som
Håkon Holien, Einar Timdal: Begerlavslekta Cladonia
Hentet fra https://artsdatabanken.no/arter/takson/44990/beskrivelse