Ampharetidae and Melinnidae
Ampharetidae and Melinnidae are families within the phylum Annelida, order Terebellida, suborder Terebellomorpha. Ampharetidae and Melinnidae are found in all the world’s oceans, and a few species of ampharetids occur in fresh water. They inhabit soft sediments from intertidal to abyssal depths but are, in general, more common in deep waters.
Systematics
Ampharetidae and Melinnidae are families within the phylum Annelida, order Terebellida, suborder Terebellomorpha. Traditionally they have been considered to be subfamilies (Ampharetinae and Melinninae) within the family Ampharetidae since they share several morphological characters, for instance, tentacles that can be retracted into the mouth, not found in other terebellomorphs. In the study by Stiller et al. (2020), based on molecular data, the former subfamily Melinninae was shown to be closer to Terebellidae, another terebellomorph family, and raised to family level.
The first Ampharetidae and Melinnidae were described from Norway. The Norwegian priest and naturalist Michael Sars described the ampharetids Amphicteis gunneri and Ampharete octocirrata in 1831 and the melinnid Melinna cristata, in 1851. Currently, worldwide, there are about 260 valid species of ampharetids and 50 valid species of melinnids. Taxonomy of the ampharetids and melinnids is complex comprising a rather high number of genera with many of them being monospecific and with insufficient diagnoses.
Ecology
Ampharetidae and Melinnidae are found in all the world’s oceans, and a few species of ampharetids occur in fresh water. They inhabit soft sediments from intertidal to abyssal depths but are, in general, more common in deep waters. Ampharetids are an important part of the fauna associated with hydrothermal vents, for example Loki Castle vent field in the Norwegian Sea (Eilertsen et al. 2017, Kongsrud et al. 2017). Both ampharetids and melinnids live in tubes which they construct of sediment particles attached to a thin layer of secreted material. Species in both families are detritovores and use their mouth tentacles to collect detritus particles.
Sampling
In Norway, most samplings of ampharetids and melinnids are done in association with environmental surveys or scientific studies, using heavy gear as grabs and sledges operated from boats. These kinds of gear collect bottom dwelling fauna together with a lot of sediment. Usually the samples are sieved, to remove as much sediment as possible, fixated in formalin or ethanol and then brought ashore to be sorted.
Sampling with heavy gear and sieving quite often damages small and soft-bodied animals. In addition, storing in a fixative result in colour fading or complete disappearance. Ampharetids and melinnids usually remain in their tubes when fixated and this might prevent the fixative to preserve them properly. The combination of these factors can make the preserved specimens looking quite different than the live worms.
In Norwegian bottom surveys, ampharetids and melinnids living in shallow waters (0–20 m) are undersampled compared to deeper ones due to use of large boats capable of carring heavy gear.
Identification
Although most species of Ampharetidae and Melinnidae in Norway have good diagnostic characters, their identification is somewhat difficult. They are small to medium sized and require high magnification to identify. Many diagnostic characters are soft body features and the preservation condition of the specimens may play a significant role in species identification. Any kind of pigmentation, including eyespots, are likely to disappear when the animals are stored in a fixative. Staining with methylene blue provides contrast to some structures and can be very helpful in observation of characters.
Ampharetids and melinnids are tube dwellers and collected specimens will usually remain in their tubes. There is a high diversity in tube-morphology. The tubes can be long or short, thin or thick, constructed of mud or sand, with different shell or plant fragments attached and with different sculpturing as annulations or veins. Some species can be identified by their tube, but this requires experience and care should be taken since different species may have similar tubes.
Most characters used for identification of ampharetids and melinnids are located in the anterior end. Here we find the prostomium, branchiae, lower lip, paleae in ampharetids and postbranchial hooks, postbranchial depression and dorsal crest in melinnids. The anterior end is usually better fixated than the rest of the worm and the posteriormost part of the abdomen is frequently completely missing.
Several different types of prostomiums can be recognized within Ampharetidae, it can, for instance, be trilobed or have longitudinal ridges and nuchal grooves. Melinnids have more or less the same type of prostomium with well-defined anterior and posterior parts, separated by a pair of deep transverse nuchal grooves. Within each type of prostomiums, there can be several variations. For instance, in species with trilobed prostomium, the middle lobe can be wide or narrow, have parallel or slanting sides, protrude higher and further than the surrounding lobe or not. Longitudinal ridges can be more or less prominent, thin or thick, be close together or further apart. The prostomium can be used to identify genera. Identification of species by the prostomium requires a lot of experience since the shape of the prostomium can change depending on the degree to which the mouth tentacles are extended, what kind of fixative has been used and how well the specimen has been fixated. For some species, the middle lobe cannot be seen when the mouth tentacles are fully extended.
The branchiae originate from several segments but appear to be clumped on segments II and III, often they are fused forming a branchial ridge across segment II. There can be two, three or four pairs of branchiae. In Ampharetidae, the branchiae can easily break off, leaving scars of attachment. In Melinnidae the branchiae are unlikely to fall off. Juveniles might not have developed all the branchiae and the fourth pair of branchiae, whenever present, appears late in development. The branchial arrangement can be hard to interpret, particular since the individual branchial scars might be hard to see.
Paleae can be found within Ampharetidae but are absent in Melinnidae. The paleae can be stout and large, but also small and unnoticeable and even missing. The tips of the paleal chaeta are important for identification, but these might break off. The number of paleal chaetae can also be a helpful character when identifying ampharetids, but care should be taken since the number is size-dependent with fewer chaetae present in the juveniles in comparison to the adults.
The postbranchial hooks present in Melinna can break off (not easily tough) leaving hardly a trace. This may lead to confusion with Melinnopsis which lacks the hooks.
In most ampharetids and melinnids, the shape of thoracic and abdominal neuropodia differs. In the thoracic type neuropodia, the uncini are located in a furrow. In the abdominal type neuropodia, the uncini are located at the margin of the neuropodia. The change from thoracic type to abdominal type happens abruptly without any transitional segments. However, the change does not necessarily correspond to the change from the thorax to the abdomen.
Many species of ampharetids and melinnids have what is referred to in literature as rudimental notopodia or dorsal papillae on the abdominal segments. These protrusions are different types of structures and, in some cases, not notopodial in origin. Often, they are ciliated, and depending on the length of the cilia, or if the cilia have fallen off, the protrusions can be more or less obvious.
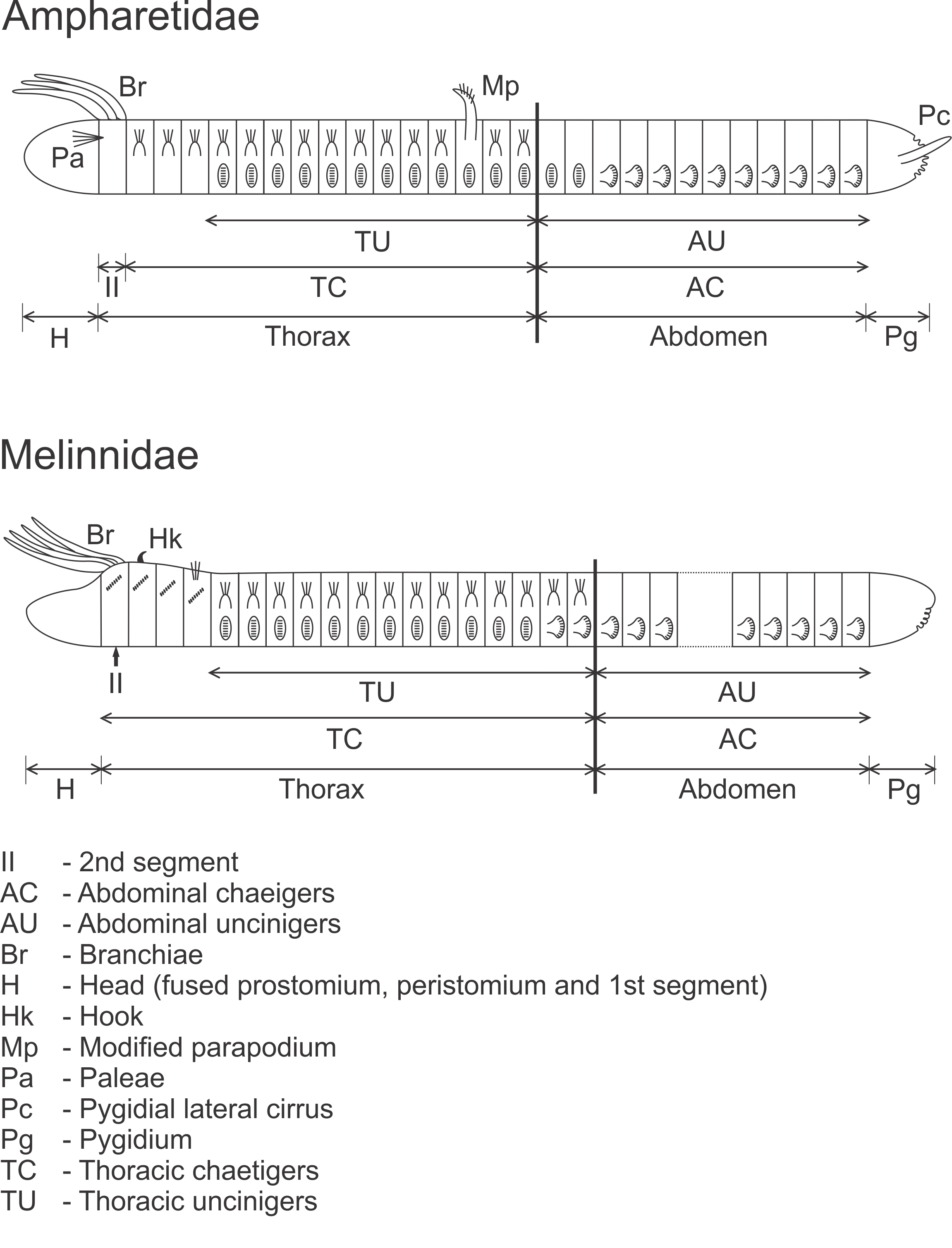
Schematic drawing of the species in Ampharetidae and Melinnidae.
Acknowledgements
We are thankful to Kennet Lundin, Gothenburg Museum of Natural History, Tina Molodtsova, Institute of Oceanology Russian Academy of Sciences, and Sergey Grebelny, Zoological Institute, Russian Academy of Sciences for help with identifying the type localities for several ampharetid species. We are also grateful to Anne Helene Solberg Tandberg, University Museum of Bergen, for help with decoding old geographical names from Svalbard.
Siden siteres som:
Alvestad T og Budaeva N (2020). Ampharetidae and Melinnidae. www.artsdatabanken.no/Pages/299547. Nedlastet <dag/måned/år>