M1 Eufotisk fast saltvannsbunn
Eufotisk fast saltvannsbunn omfatter fast fjell og stabile blokker med permanente samfunn av flerårige alge- og dyrearter, i beltet mellom normalt fjæremål, det vil si nivået som vanligvis konstant fuktes av havvann (upper infralitoral zone), gjennom tareskogsbeltet (lower infralitoral zone) og rødalgebeltet (upper circalitoral zone) ned til kompensasjonsdypet, der lysinnstrålingen er så lav [< 0.3–1(–10) % av lysinnstrålingen på ei åpen havoverflate] at planteproduksjonen er mindre enn nedbrytningen og alger ikke lenger forekommer regelmessig (netto primærproduksjon går fra å være positiv til å bli negativ).

Hovedtypen inneholder både skrånende fast fjell («knaus») og loddrette vegger. Knausene domineres av algesamfunn; på steder med optimale forhold (moderat bølge- og strømpåvirkning, ikke utsatt for direkte bølgeslag eller tørrlegging, gode lysforhold og normal salinitet) finner vi langs hele kysten velutviklete tareskoger [dominert av stortare (Laminaria hyperborea) og sukkertare (Saccharina latissima)]. Mot mindre gunstige kombinasjoner av leveforhold overtar andre arter eller artsgrupper; grønnalger på steder med redusert saltholdighet og/eller liten vannbevegelse og rødalger i dybdebeltet under tareskogsbeltet. Bergvegger og knauser som er svært sterkt eksponert for bølger eller tidevannsstrømmer har spesielle organismesamfunn som er dominert av heterotrofe organismer som dødningehånd (Alcyonium digitatum) og ulike sekkdyr-arter (Tunicata: Ascidiacea).
Tareskog. Med tareskog menes et sammenhengende område dominert av tare-arter (Laminaria spp., Saccharina latissima), med areal større enn 100 m2 og bredde større enn 5 m. Tareskogsbunn forekommer i normalsalt vann (> ca. 30 ‰), først og fremst i dybdebeltet 2–15 m under fjæremålet, på fast bunn som er beskyttet til nokså eksponert. Tareskog finnes langs hele norskekysten. Den mengdemessig viktigste tare-arten er stortare (Laminaria hyperborea), som danner tette «skoger» av 1–3 m høye planter. Hver tareplante består av en stiv, lang stilk som bærer ett blad. Også sukkertare (Saccharina latissima) og andre tarearter som fingertare (Laminaria digitata), butare (Alaria esculenta) og draugtare (Saccorhiza polyschides) kan danne tette bestander«skoger», og tareskogene kan også være «blandingsskoger» der to eller flere arter dominerer sammen. Sukkertaren har kortere stilk enn stortare (20–30 cm lang). Sukkertarestilken bærer et blad som kan bli over 2 m langt. Sukkertareskogene ser derfor mer ut som store avlange blader som dekker bunnen. Stortare og sukkertare har ulike krav til vannbevegelsesenergi og dominanstypene avløser hverandre derfor langs LKM vannpåvirkningsintensitet (VF). Stortare finnes der det er god vannbevegelse forårsaket av strøm og bølger (VF∙ef, nokså eksponert), mens sukkertare trives under mer beskyttete forhold og typisk forekommer i skjærgården der den stopper litt innenfor fjordmunningene (VF∙cd). Stortaredominerte tareskoger finnes best utviklet fra Lindesnes til Finnmark, og er aller frodigst i Midt-Norge. Tareartene og mange assosierte arter er avhengig av brukbar lystilgang og tareskog finnes derfor fra grensa mellom saltvannssystemer og fjærebeltesystemer til et sted mellom 15 og 30 meters dyp, avhengig av vannkvaliteten. I Nord-Norge har tareskog blitt observert ned til 37 m – det er lyset som begrenser nedre voksedyp.
Tareskogene er blant de mest produktive økosystemene på jorda. Produksjonsmålinger fra Midt-Norge viser en årlig produksjon opp til 3 kg karbon pr. m2 (Abdullah & Fredriksen 2004). Dette tallet er oppsiktsvekkende høyt og tilsvarer produksjon av ca. 25 kg plantemateriale pr. m2 og år. Taren er vert for produktive samfunn av påvekstorganismer (blant annet rødalger som vokser som epifytter på tarens stilk). Totalproduksjonen i tareskogen er større enn på dyrket mark på land. Det potensielle arealet av (tang- og) tareskoger langs norskekysten er anslått til å være på størrelse med arealet dyrket mark på det norske fastlandet, så tareskogene har stor betydning for den totale produksjonen i kystnære farvann (enda større ville den vært dersom den ikke hadde blitt redusert av nedslamming og nedbeiting).
Tareskog har mange fellestrekk med skogsmark på land. Tareskogen er et tredimensjonalt system med stort nisjemangfold og dermed stort artsmangfold både av planter og dyr (se Christie et al. 2003). Planteartsmangfoldet er i sin tur med på å øke mangfoldet av dyrehabitater, og dyr (mest krepsdyr og snegler) utnytter hele det tredimensjonale systemet. Smådyr blir utnyttet av et stort mangfold av fiskearter, der særlig unge stadier av torsk (Gadus morhua), sei (Pollachius virens) og lyr (Pollachius pollachius) er tallrike, men der også krabbe-arter, hummer-arter og sjøfugl utnytter det rike dyrelivet. Tareskogen stabiliseres blant annet gjennom biologiske reguleringsmekanismer som involverer naturtyper over og under tareskogsbeltet. Snegler og krepsdyr i tette bestander lever stort sett av overskuddet fra planteproduksjonen, og fiskene sørger for å regulere bløtdyr- og leddyrtettheten slik at nedbeiting av taren, som både er leveområde og primærprodusent i denne natursystem-hovedtypen, unngås.
Flere ytre faktorer bidrar imidlertid til tidvis ustabilitet og stor dynamikk i tareskogene. Tareskoger kan utsettes for store ødeleggelser under stormer, men liksom i skogsmark etter vindfall rekrutteres nye tareplanter raskt. De dominerende tareartene har stor konkurransekraft og hindrer vanligvis andre arter i å dominere. Tareskogens artsinventar (både av planter og dyr) er tilpasset stadige omskiftinger, og gjenetablert tareskog blir raskt kolonisert av en rekke organismer slik at en «moden»tareskog er etablert allerede etter 4–5 år. Taretråling forårsaker noen av de samme skadene som storm, men tareskogen regenererer etter tråling. Det har vært antydet at høsting hvert femte år kan være bærekraftig, men det er tvilsomt om den gjenetablerte tareskogen blir «økologisk moden» (det vil si at den regenererer en fullstendig eller nesten fullstendig artssammensetning) dersom den tråles igjen allerede etter fem år; det er mulig at gjentatt trålvirksomhet vil ha økologiske effekter av lengre varighet.
Bortfall av sukkertareskog i Skagerrak og langs vestlandskysten, og nedbeiting av stortareskog langs kysten av Midt- og Nord-Norge, forårsaket av drøbak-kråkebollen (den grønne kråkebollen, Strongylocentrotus droebachiensis) og rød kråkebolle (Echinus esculentus) fører til sterk ubalanse i tareskogsøkosystemet fordi det første leddet i næringskjeden (taren) fjernes. Når taren forsvinner, forsvinner også systemets tredimensjonale struktur og systemets funksjon kan kollapse. Årsakssammenhengene bak disse fenomenene er ikke fullstendig kjent. Det pågår imidlertid forskning som skal belyse betydningen av endringer i temperaturen, tilførsel av næringssalter, nedslamming og forekomst av algematter, som mulige årsaker til reduksjon i sukkertare-dominerte tareskoger på Skagerrakkysten. En mulig forklaring er at sukkertaren i Skagerrak dør som følge av økt temperatur og økt næringssalttilførsel, og at økende tildekking av bunnen (som følge av nedslamming og etablering av hurtigvoksende trådformete alger) hindrer reetablering av nye tareplanter slik at tareskogen ikke kommer tilbake. Både tang og tare er avhengig av fast bunn fordi disse organismene bare fester seg til «fjellbunn». Arealet av sukkertareskog i tilbakegang er kartlagt og berører strekningen fra Hvaler (Østfold) til Lindesnes (Vest-Agder) og videre nordover langs vestlandskysten til Møre (Moy et al. 2007). Det er bare sukkertareskogen som er sterkt berørt, mens stortaredominert tareskog langs den samme kystlinjen ikke synes å være utsatt i samme grad. Dette skyldes sannsynligvis at stortare finnes på lokaliteter med større vannbevegelse, der faren for nedslamming og tilgroing er mindre.
Nedbeiting forårsaket av kråkeboller har langt større omfang enn nedslamming og tilgroing. Fenomenet som først ble oppdaget for ca. 35 år siden, da den grønne kråkebollen (Strongylocentrotus droebachiensis) ble observert i store tettheter og store deler av tareskogen fra og med Trøndelag til og med Finnmark ble beitet ned til en tilstand som kan karakteriseres som en «marin ørken». I dag er anslagsvis 90 % av tareskogen i Finnmark nedbeitet, og grove estimater antyder at i overkant av 2 000 km2 med tareskog er i nedbeitet tilstand. Sørgrensa for det nedbeitete området flytter seg nordover og befant seg i 2007 i Brønnøysundområdet (Norderhaug & Christie 2007). Beregninger indikerer at kråkebollebeiting er årsak til et årlig tap av potensiell tareproduksjon på over 20 millioner tonn, det vil si over 100 ganger mer enn det som fanges av taretrålere. Verdien av dette tapet (av tareproduksjon og fiskeproduksjon) kommer opp i milliardbeløp. Årsaken til kråkebolleoppblomstringen for 35 år siden har forblitt ukjent, men tilsvarende fenomener andre steder i verden kan gi hint om mulige forklaringer. På stillehavskysten av Nord-Amerika førte overbeskatning av havoter til et liknende problem, mens modellberegninger har indikert at overfiske av torsk er den mest sannsynlige årsaken til tilsvarende fenomen på atlanterhavskysten (se for eksempel Steneck et al. 2002, 2004). På samme vis kan redusert predatorkontroll (fiskebestanden bidrar normalt til kontroll av størrelsen på beitedyrpopulasjonene) være en mulig årsak til overbeiting av den norske tareskogen. I motsetning til mange andre dyr klarer kråkebollene å overleve i høye tettheter på den nedbeita bunnen ved å beite på det som måtte finnes av mat der. Samtidig som bunnen holdes fri for tare, holdes også alle potensielle fiender og konkurrenter (som normalt holder til i tareskogen) borte. Alt dette stabiliserer og forlenger tilstanden av overbeite. Dersom kråkebolletettheten av en eller annen grunn blir tilstrekkelig lav, utvikles ny tareskog. Nedbeiting synes derfor, i hvert fall på kort sikt, ikke å føre til irreversible endringer i tareskogsøkosystemet. I framtida kan også klimaendringer komme til å påvirke balansen mellom tare og den grønne kråkebollen, fordi sistnevnte er en kaldtvannsart.
Definisjonsgrunnlag og avgrensning
Variasjon
Så vel populærbeskrivelser av variasjonen på hardbunn i det eufotiske beltet (f.eks. Køie & Kristiansen 2000, Fredriksen & Throndsen 2001) som vitenskapelige typesystemer som det britiske MarLIN (Connor et al. 2004), identifiserer vannpåvirkningsintensitet (VF), dybderelatert lyssvekking (DL) og marin salinitet (SA) som de viktigste LKM for variasjon i artssammensetning på eufotisk fast saltvannsbunn. I tillegg pekes på betydelige forskjeller i artssammensetning mellom bergvegger, med for sterk helningsbetinget forstyrrelsesintensitet (HF) til at tang- og taresunnfunn finnes (sannsynligvis grunnet i etableringsproblemer), og knauser. Liksom for T1 Nakent berg synes det som om det går et relativt skarpt skille i artssammensetning omkring 70–80 ° helning (K.M. Norderhaug, pers. medd.). Dette er tydelig vist for et undersøkelsesområde på Svalbard av Jørgensen & Gulliksen (2001). Også innenfor vertikale vegger er det klar dybdesonering (Gulliksen 1978, Jørgensen & Gulliksen 2001). Dybdebeltene er klart definert med hensyn til artssammensetningen, men inntar ikke et fast dybdeintervall, målt i m. Beltene «forskyves’ mot grunnere vann på steder med redusert lys, f.eks. ligger beltegrensene dypere på skyggefulle, nordvendte fjordsider enn på med åpent plasserte, sørvendte fjordsider.
På steder med sterk vannpåvirkningsintensitet (VF) er det dessuten forskjeller i artssammensetning mellom steder der den sterke vannpåvirkningsintensiteten skyldes sterke tidevannsstrømmer og der den skyldes sterke bølger (Connor et al. 2004). Dette fanges opp av LKM Vannpåvirkningsregime (VR).
Stabile grove sedimenter [først og fremst blokker; dominerende kornstørrelsesklasse (S1) S1∙a] har også en artssammensetning dominert av påvekstorganismer slik som bergknausene, men får i tillegg nye nisjer på grunn av den ujevne overflata med hulrom, blokkundersider etc. Her lever fisker, krepsdyr etc. Det er usikkert hvor stor endringen i artssammensetning i forhold til bergknauser er, men inntil det foreligger mer informasjon er S1 Dominerende kornstørrelsesklasse lagt inn som en tLKM. Den videre oppdelingen av stabil marin blokkmark (S1∙a) er tentativ og tar utgangspunkt i hoveddiagrammet for hovedtypen. Eksplisitt kunnskap mangler.
Det er stor naturlig dynamikk i marine fastbunnssystemers artssammensetning, til dels relatert til variasjon i miljøforholdene, som vist av Beuchel et al. (2006), Kortsch et al. (2013) og Ballantine (2013) ved repetert fotografering av permanente fastbunnsstasjoner på Svalbard. Denne dynamikken kan bare delvis forklares av (variasjon i) miljøforholdene; f.eks. fant Beuchel et al. (2006) at 45 % av mellomårsvariasjonen på en overvåkingsstasjon i Kongsfjorden som ble fulgt fra 1980 til 2003 kunne forklares av miljøforhold relatert til klimavariasjon (uttrykt som NAO; «The North Atlantic Oscillation»).Kortsch et al. (2012) dokumenterer imidlertid at plutselige endringer i makrobentos-samfunn kan inntreffe; i Kongsfjorden og Smeerenburgfjorden fant det på 2000-tallet sted en rask økning i dekningen av de boreale makroalgene kjerringhår (Desmarestia spp.), eikeving (Phycodrys rubens) og bladtare (Saccorhiza dermatodea). Økningen i dekning kunne være inntil 8× på ett år! Raske kolonisering ble også observert på lavabunn på Jan Mayen etter at et vulkanutbrudd la ny bunn åpen for kolonisering; allerede etter 8 år var faunasammensetningen omtrent lik på lavabunn og på «gammel bunn’ (Gulliksen et al. 1980). Dette indikerer at rask og stor dynamikk er normalt på fast eufotisk bunn, og er et viktig signal om at gradientlengdene langs de viktige LKM kanskje er mindre enn man først kan være fristet til å anta.
Det finnes ikke undersøkelser som kvantifiserer variasjonen i artssammensetning langs de LKM som er identifisert som viktige. Til grunn for trinninndelingen er derfor lagt at godt karakteriserte «samfunn» som avløser hverandre langs de aktuelle LKM er akseptert som trinn. VF er dermed delt i to sentrale trinn for de tareskogsdominanstypene med ett trinn på hver side. Likeledes er DL delt i de tre trinnene som vanligvis er beskrevet i litteraturen. Liksom langs andre viktige LKM, er det lagt til grunn at ekstreme forhold langs én gradient har en sterk tendens til å «overstyre» og jamne ut variasjon langs andre. Følgelig er grunntypeskjemaene forenklet slik at bare kombinasjoner av LKM-trinn som er kjent for å ha, eller med stor sannsynlighet har, en distinkt artssammensetning, er opprettholdt som grunntyper.
Tredelingen av marin salinitet (SA) grunner seg på observasjoner omkring utløpet av Glomma (@ref) og i Sandnesfjorden (T. Kroglund, @sjekk) av hvor langs denne LKM at ulike algearter kommer inn, og indikasjoner på at det er viktige endringer i artssammensetningen ved saliniteter omkring 18 og 5 ‰ (F. Moy & E. Oug, pers. medd.)
Det er ikke klart hvor stor den systematiske forskjellen i artssammensetning mellom tidevannsstrøm- og bølgepåvirkete sterkt eksponert eufotisk fast bunn er. VF er derfor tentativt inkludert som uLKM inntil det eventuelt blir vist at det grunnlag for å oppgradere den til tLKM.
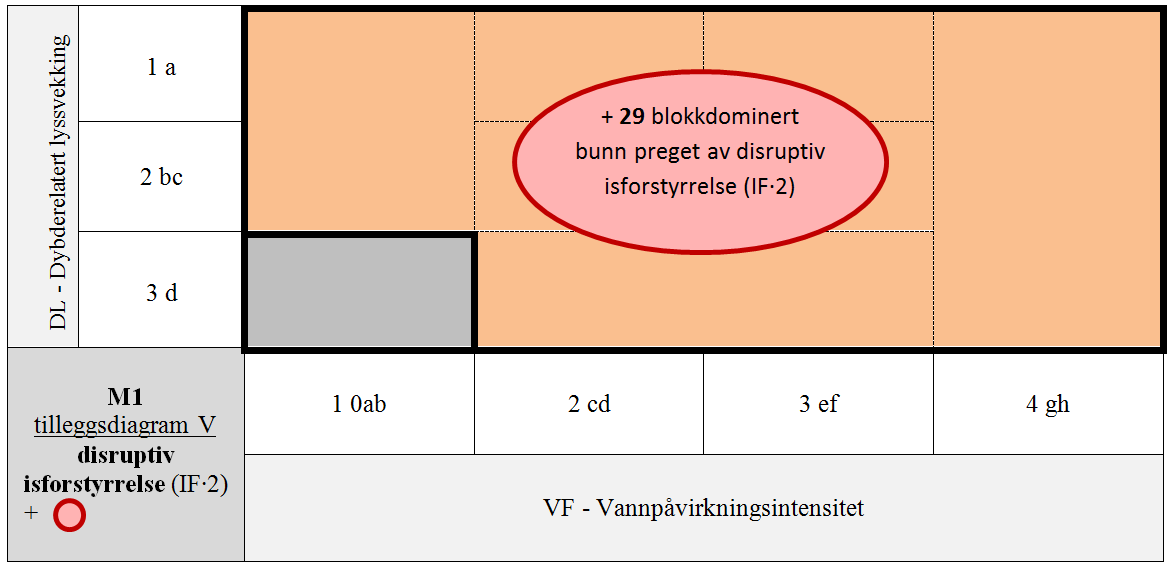
I Arktis forårsaker disruptiv isskuring (IF∙¤) stedvis mer eller mindre naken bunn helt ned til 6–8 m dyp (Gulliksen 1979). Tidvis fullstendig tap av arter og at samfunn av flerårige organismer ikke opprettholdes over lengre tid betinger, liksom disruptiv sedimentasjonsbasert forstyrrelse (SE∙¤) og vindutsatthet (VI∙¤) at isbetinget forstyrrelse (IF∙¤) inkluderes som grunntyper i M1. Isforstyrrelsesvirkningen er uavhengig av hvilke arter som dominerer, slik at det ikke er finnes noen gradient i artssammensetning mellom intakt fast bunn (IF∙0ab) og bunn med disruptiv isforstyrrelse (IF∙¤). Til grunn for bare å skille ut én grunntype for eufotisk fast saltvannsbunn preget av disruptiv forstyrrelse ligger at denne LKM overstyrer variasjon langs alle andre LKM.
Raspreget fast eufotisk saltvannsbunn forekommer mange steder langs kysten der rasmarker går rett i havet. Fordi suksesjonen på naken marin fast bunn går mye raskere enn på nakent bergsubstrat på land, må rasintensiteten være desto sterkere for å gi utslag på artssammensetningen. Artssammensetningen på marin rasmark er dårlig kjent, men marin faggruppe konkluderte at høy rasfrekvens sannsynligvis gir seg utslag i artssammensetningen, i hvert fall i form av redusert artsrikdom. Som en konservativ løsning er rasutsatthet (RU) inkludert i beskrivelsessystemet for M1 som en uLKM.
Kunnskapsbehov
Sammenstilling av generaliserte artslistedatasett (fortrinnsvis flere, fra ulike regioner) for å teste den framlagte naturtypeinndelingshypotesen med henblikk på gradientlengdeberegning for alle foreslåtte hLKM, tLKM og uLKM.
Analyse av artslistedata fra MarLIN, med stasjoner og typer fordelt på naturtypekategorier gitt av framlegget til typeinndeling i NiN 2.
Grunntypeinndeling
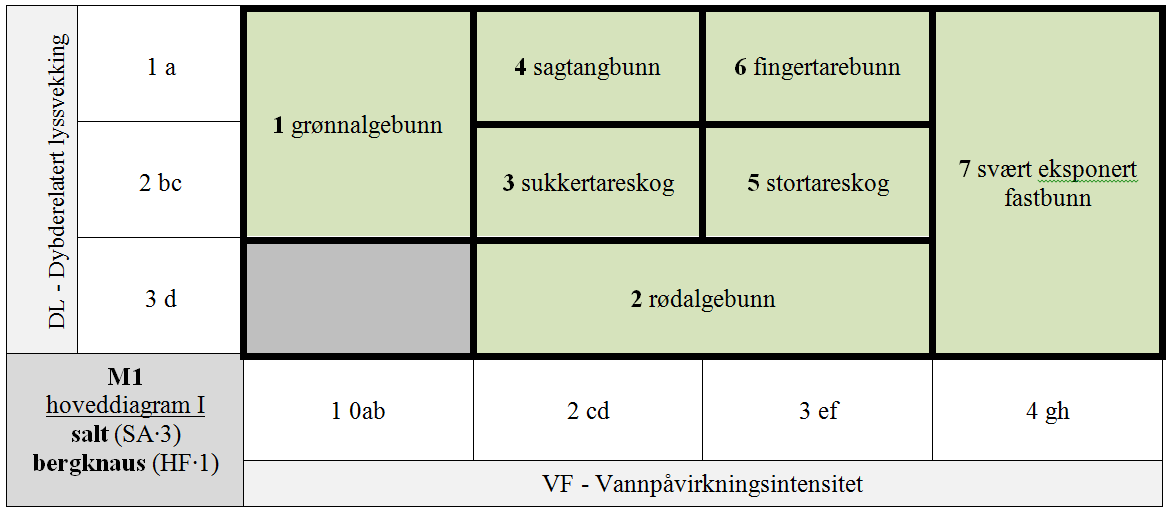
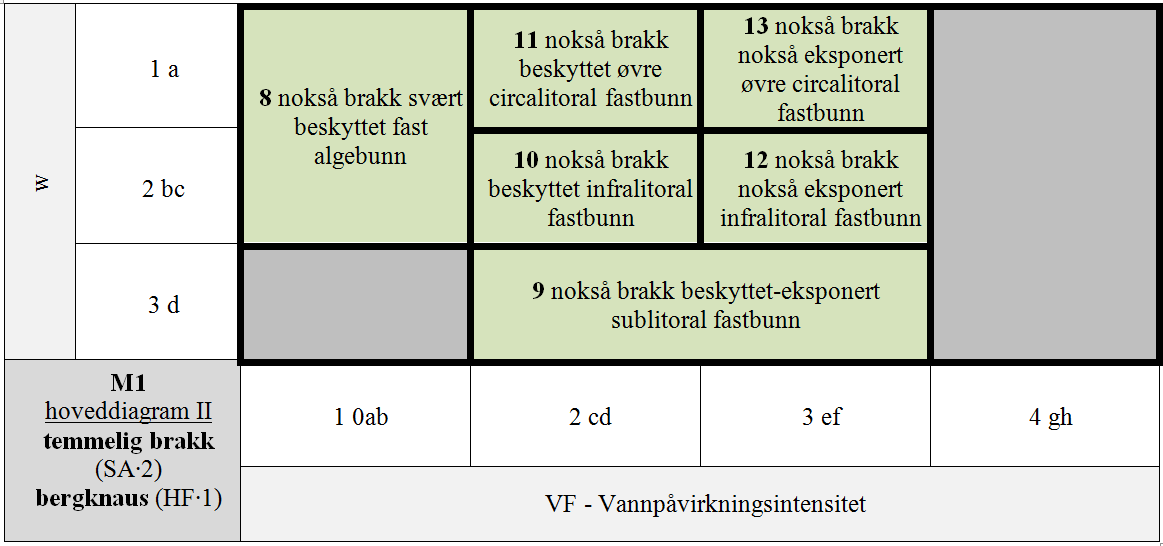
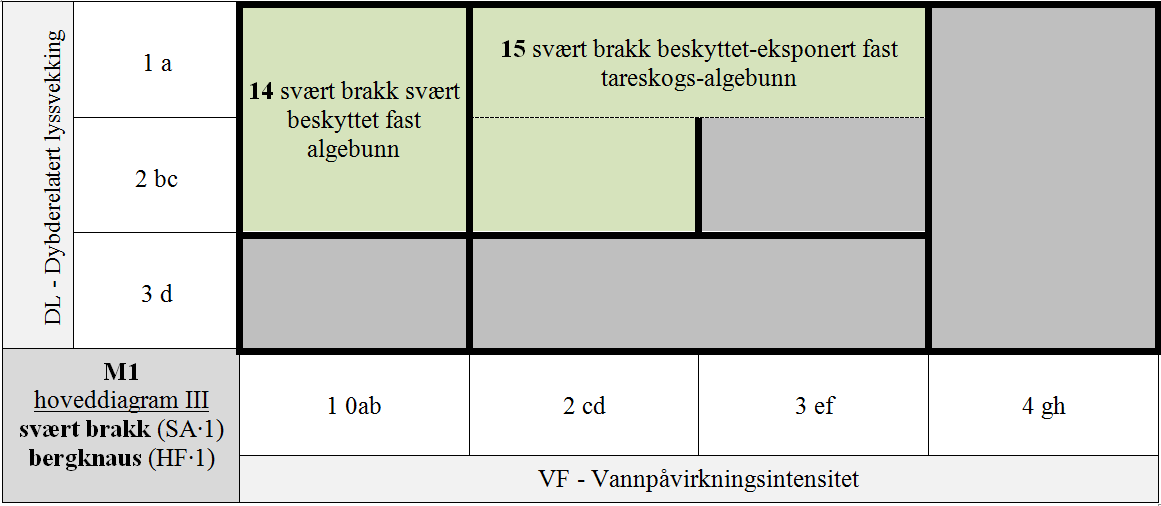
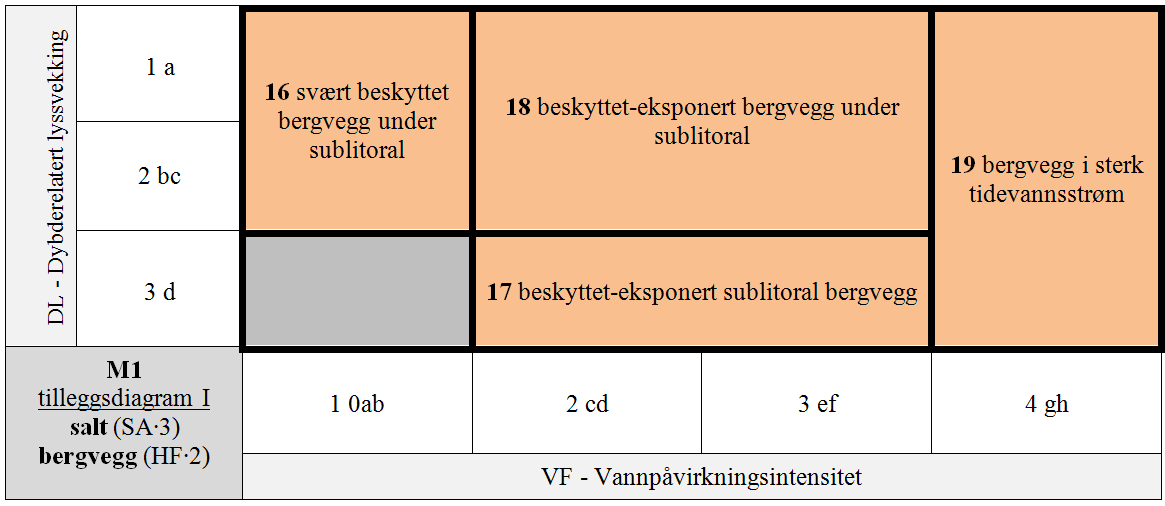
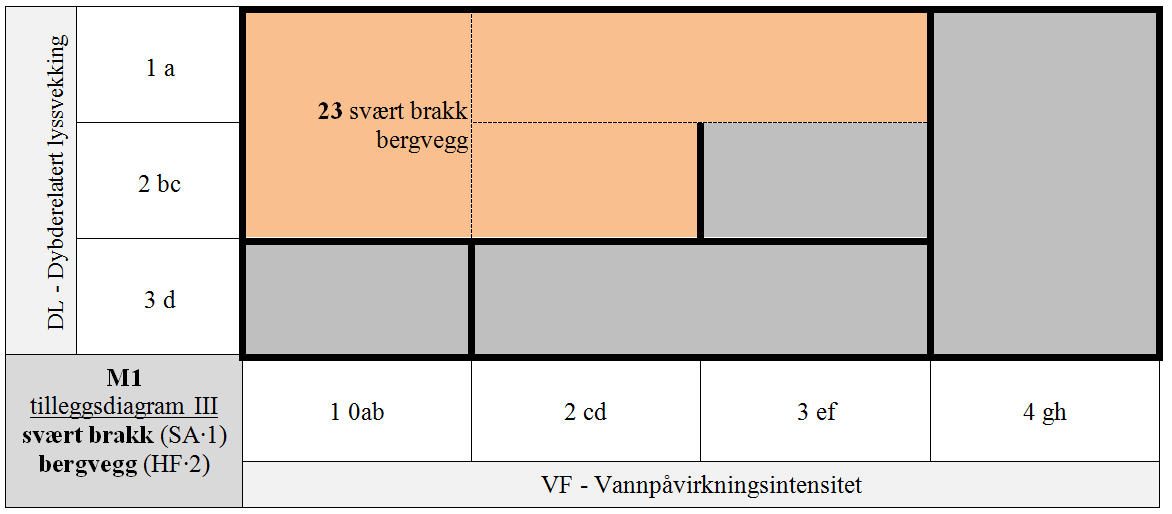
M1 er delt i 7 GT for kombinasjoner av hLKM 4 VF × 3 DL for SA∙3 & HF∙1 & S1∙A (normalsalte bergknauser) som antas å ha innbyrdes betydelig forskjellig artssammensetning, + 8 GT for kombinasjoner VF × DL med redusert saltholdighet (SA∙2 og SA∙1), + 4 GT for kombinasjoner VF × DL for HF∙2 & SA∙3 & S1∙A (normalsalte bergvegger), + 4 GT for spesielle kombinasjoner av VF × DL med redusert saltholdighet (SA∙2 og SA∙1) for HF∙2, + 4 GT for spesielle kombinasjoner av VF × DL på stabiliserte blokker (S1∙B) for HF∙1 & SA∙3, + 1 GT for den spesielle kombinasjonen av stabiliserte blokker (S1∙B) og redusert saltholdighet (SA∙2), + 1 GT for IF∙2 uavhengig av trinn langs andre LKM.
- M1-1 stille til temmelig beskyttet fast algebunn
- M1-2 litt beskyttet-svakt eksponert sublitoral fastbunn
- M1-3 beskyttet infralitoral fastbunn
- M1-4 beskyttet øvre circalitoral fastbunn
- M1-5 nokså eksponert infralitoral fastbunn
- M1-6 nokså eksponert øvre circalitoral fastbunn
- M1-7 svært eksponert eufotisk fastbunn
- M1-8 nokså brakk svært beskyttet fast algebunn
- M1-9 nokså brakk beskyttet-eksponert sublitoral fastbunn
- M1-10 nokså brakk beskyttet infralitoral fastbunn
- M1-11 nokså brakk beskyttet øvre circalitoral fastbunn
- M1-12 nokså brakk nokså eksponert infralitoral fastbunn
- M1-13 nokså brakk nokså eksponert øvre circalitoral fastbunn
- M1-14 svært brakk svært beskyttet fast algebunn
- M1-15 svært brakk beskyttet-eksponert fast tareskogs-algebunn
- M1-16 svært beskyttet bergvegg under sublitoral
- M1-17 beskyttet-eksponert sublitoral bergvegg
- M1-18 beskyttet-eksponert bergvegg under sublitoral
- M1-19 svært eksponert bergvegg
- M1-20 svært beskyttet nokså brakk bergvegg
- M1-21 beskyttet-eksponert nokså brakk bergvegg i sublitoral
- M1-22 beskyttet-eksponert nokså brakk bergvegg under sublitoral
- M1-23 svært brakk bergvegg
- M1-24 svært beskyttet blokkdominert bunn under sublitoral
- M1-25 beskyttet-eksponert blokkdominert bunn i sublitoral
- M1-26 beskyttet-eksponert blokkdominert bunn under sublitoral
- M1-27 svært eksponert blokkdominert bunn
- M1-28 beskyttet-eksponert nokså brakk blokkdominert bunn under sublitoral
- M1-29 blokkdominert bunn preget av disruptiv isforstyrrelse